| <<< Előző cikkhez | Következő cikkhez >>> |
| MAGYAR TUDOMÁNY cikke | Téma : Őstörténetünkhöz | |
Genetika és (magyar) őstörténet | ||
Archeogenetikai vizsgálatok a Kárpát-medence 10. századi népességénA cikk a Magyar Tudomány,
2008/10 számában jelent meg.
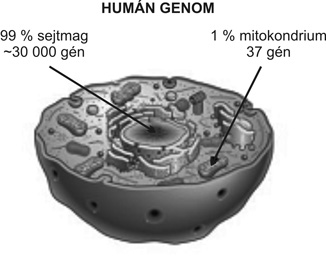
Az ember külső és belső tulajdonságait az örökítőanyag, a DNS határozza meg. Ugyanez a molekula az emberiség eredetére, de ezen belül az egyes népcsoportok eredetéről és rokonsági viszonyairól is hordoz információkat, ugyanakkor közvetlen rokonsági kapcsolatok kimutatására is alkalmas. Az ember teljes genetikai információja, a genom közel 99 %-a a sejtmagban, kromoszómákba csomagolva, míg a maradék 1 %-a a mitokondriumokban található. Mai ismereteink szerint a mitokondriális DNS 37 gént hordoz, míg a sejtmagi, kromoszómákba tömörült ún. nukleáris géneknek a száma kb. 25–30 ezer (1. ábra). A sejtmagi géneket mindkét szülő egyenlő mértékben hagyományozza ránk, ráadásul az egyedfejlődés egy bizonyos stádiumában ezek keverednek, rekombinálódnak is egymással. A legutolsó nemzedékben két nukleáris felmenőnk van, egy generációval „feljebb” már négy nukleáris elődöt találunk. Húsz emberöltőre visszamenően, átlagosan huszonöt éves generációs idővel számolva, több mint egymillió azon elődök száma, akik hozzájárultak sejtmagbéli génjeinkhez. Jelenleg az emberiség történetének különféle egyedi változásait kiolvasni a sejtmag génjeiből és géndarabjaiból reménytelenül bonyolultnak tűnik. Populációk rokonsági viszonyainak tanulmányozásához ezért célszerű olyan bélyegeket vizsgálni, amelyekben nem keveredik az anyai és az apai információ az egymást követő nemzedékekben. Ellentétben a nukleáris DNS döntő hányadával, a mitokondriális DNS öröklődése lineáris, vagyis a genetikai információ forrása nemzedékenként csak az anya, illetve az anyai ági öröklési vonal. Az Y-kromoszóma egy bizonyos szakasza pedig változatlanul öröklődik apáról fiúra. Régi és mai populációk rokonsági viszonyának megértéséhez, a régi populációk mozgásának meghatározásához egyik lehetséges módszer e populációk genetikai jellegzetességeinek összevetése. Ennek feltétele, hogy archaikus mintákban is kimutathatók és tipizálhatók legyenek az örökítőanyag kutatási céloknak megfelelő szakaszai. A régészeti mintákban a DNS az emberi és állati testet felépítő szöveti elemek közül a csontokban és a fogazatban vizsgálható, ahol a konzerválódás és a DNS-tartalom szerkezeti megőrződése elsősorban a temetkezési hely mikrokörnyezeti adottságainak függvénye. A Kárpát-medence többnyire savas talajközegében lévő maradványokban általában csekély mennyiségű és rossz minőségű DNS mutatható ki. Jellemzője az erős töredezettség, oxidatív és hidrolitikus bázismódosulások, a talajból és a biológiai anyag degradációjából származó, kapcsolódó vegyületek jelenléte. Vizsgálatainkhoz a DNS-t csöves csontokból nyertük. Az izolált DNS vizsgálni kívánt szakaszát ún. polimeráz-láncreakcióval (PCR) felsokszoroztuk, majd a reakció sikerességét és hitelességét gél-elektroforézissel ellenőriztük. A felsokszorozott DNS-régiót végül a bázissorrend megállapítása érdekében szekvenáltuk (2. ábra).
I. A mitokondriális DNS öröklődési vonal vizsgálata
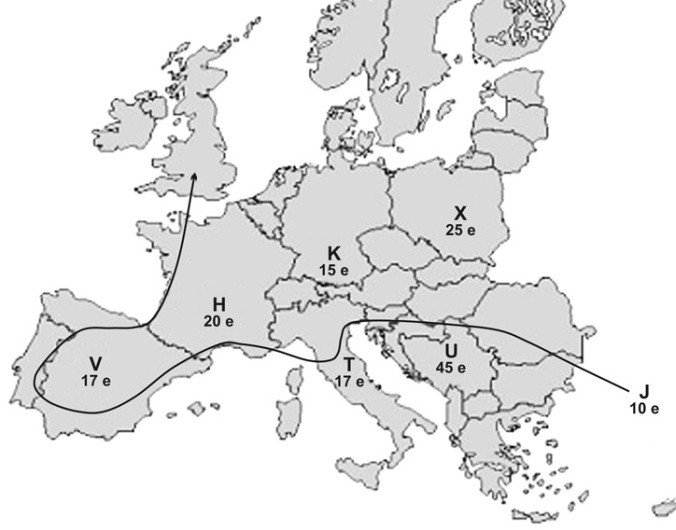
A mitokondriumok feladata, hogy segítsék az oxigén felhasználását a sejt energiatermelésében. Minél tevékenyebb a sejt, annál több mitokondriumot tartalmaz. A DNS-változások, -mutációk mind a mitokondriumban, mind a sejtmagban spontán jönnek létre, általában egyszerű másolási hibák, amelyek a sejtekben működő hibajavító mechanizmusok ellenőrző rendszerén észrevétlenül átcsúsznak. A mitokondriális DNS-ben van egy kb. 1200 bázispáros nem kódoló – az ún. hipervariábilis (HVS) – régió, ahol a bekövetkezett mutációk nem járnak az organizmus számára káros következményekkel, így az itt kialakult mutációk gyorsan rögzülnek, és aránylag gyorsan elterjedhetnek a populációban. Mivel a szakaszon kb. minden tízezredik évben rögzül egy mutáció, ezért ez molekuláris evolúciós óraként is használható. Az egyedi szekvenciák egymástól eltérő pozícióik (mutációik) alapján egy-egy haplotípust határoznak meg. Azok a szekvenciák megegyező haplotípusúak, amelyek ugyanazokat a mutációkat tartalmazzák. A haplotípusokat haplocsoportokba soroljuk egyes ősi, azonos mutációik alapján. Azok a mutációk, amelyek egy haplocsoportot jellemeztek hosszú idővel ezelőtt, a filogenézis során csak egyszer alakultak ki, és többségükben jellegzetes földrajzi eloszlást mutatnak. Többezer mitokondriális DNS-szekvencia összehasonlításával a kutatók arra a következtetésre jutottak, hogy az összes ma élő ember anyai származásvonalának gyökerénél egyetlen nő áll. Ezt a gyökérszekvenciát elnevezték „mitokondriális Évának”, akinek leszármazottai aztán szétterjedtek az afrikai kontinensen. Az anatómiailag modern Homo sapiens képviselői kb. 75–55 ezer éve hagyták el Afrikát a közel-keleti és arábiai területeken keresztül (ún. Out of Africa-elmélet) Közép-Ázsia felé. E vándorlás közben, földrajzilag a közel-keleti és közép-ázsiai területek közötti zónában két fontos alap-haplocsoportra oszlottak (M és N), amelyekből az összes nem afrikai eredetű haplocsoport származtatható. Az M haplocsoport leszármazottai keleti és délkeleti irányba rajzottak szét, benépesítve Eurázsia keleti területeit, Ausztráliát és Amerikát. Az N haplocsoport utódnemzedékei a paleolitikum és a neolitikum idején Európát, illetve részben Ázsiát népesítették be, több hullámban, a Közel-Keleten keresztül. Emellett a neolitikus időszakban feltételeznek egy, az Uralból induló, és Európa északi részét érintő vándorlási vonalat. A recens európai népesség 95 %-a hét fő genetikai nyalábba, haplocsoportba sorolható (3. ábra). A hat csoport a paleolitikum idején alakult ki Európa különböző területein, míg a legfiatalabb, valószínűleg a Közel-Keleten kialakult haplocsoport (J), mintegy tízezer évvel ezelőtt, a neolitikus népmozgások kapcsán került az európai kontinensre.
Pályázati munkánk első időszaka alatt a honfoglaló magyarság mtDNS-mintázatának leírásához 79 leletet vizsgáltunk meg. Először minden, a régészek által kiválasztott temetkezési helyről három leletet analizáltunk, hogy megállapítsuk, az adott mikrokörnyezeti viszonyok között maradt-e a csontban kimutatható mennyiségű és minőségű archaikus DNS. A 79 csontlelet közül 42 esetben tudtuk az ősi DNS jelenlétét kimutatni. Vizsgálataink során a mitokondriális DNS hipervariábilis I. régiójának egy 360 bázispáros szakaszát tanulmányoztuk két rövidebb, részben átfedő szakasz illesztésével. A polimorf pozícióik alapján az egyes szekvenciákat haplocsoportokba soroltuk. A 42 lelet közül 35 esetben tudtuk kimutatni a megfelelő hosszúságú DNS-szakaszt, 26 minta esetében a kódoló régió egyes haplocsoport-specifikus polimorf pozícióinak meghatározásával sikerült a haplocsoportba sorolás. A második pályázati időszak alatt összesen 57 db. 10. századi csontminta vizsgálatát végeztük el. 43 minta esetében sikerült a mitokondriális analízishez szükséges hosszúságú DNS-szakaszt felsokszoroznunk, illetve a haplokategorizáláshoz szükséges polimorf pozíciókat meghatároznunk. Kilenc csontmintánál sikerült ugyan archaikus DNS-t izolálnunk, de a mutációs pontok meghatározása tekintetében egyelőre csak részeredmények állnak rendelkezésünkre, ami e minták analíziséhez nem elegendő.
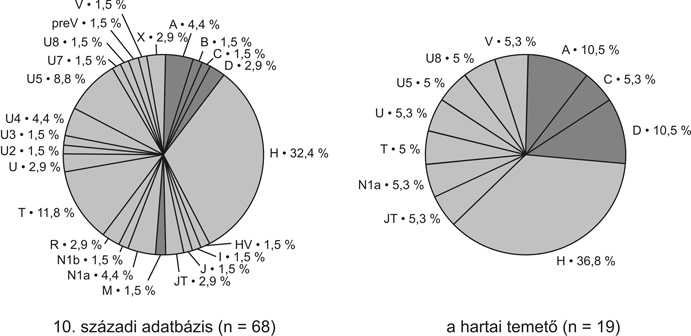
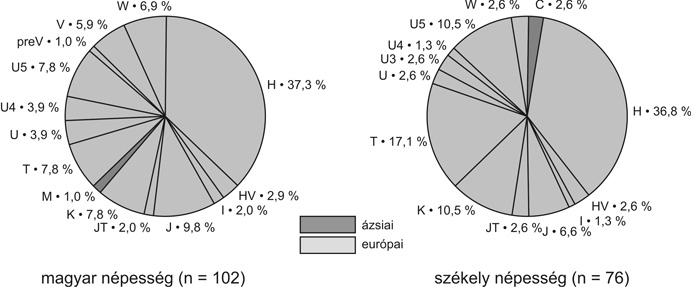
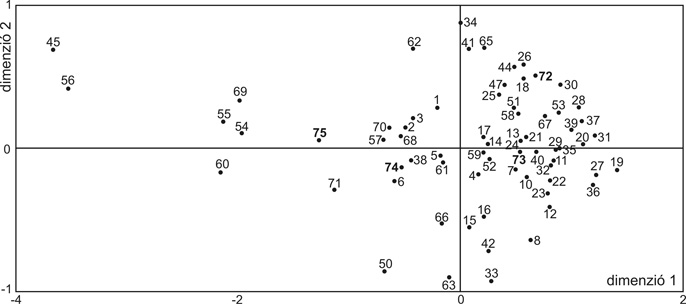
Eddigi kutatásaink során összesen 68 honfoglalás kori csontmintából kaptunk értékelhető eredményt. Ezek 24 nagyobb, európai, illetve ázsiai haplocsoportba tartoztak. (4. ábra). Az összes szekvencia 11,8 %-a kifejezetten ázsiai haplocsoportba (A, B, C, D, M) sorolható, míg 4,4 % az N1a haplocsoport ázsiai ágán található. Így összesen a minták 16,2 %-a képvisel ázsiai vonalat, míg a maradék 83,8 % európai haplocsoportokba tartozik. A minták 32,4 %-a az Európában leggyakrabban – általában 40–60 %-ban – előforduló H haplocsoportot képviseli. A második leggyakoribb az U haplocsoport 22,1 %-os gyakorisággal. Ezen belül 13,2 % tartozik az Északkelet-Európában, a volga–uráli népeknél és Nyugat-Szibériában gyakori U4 és U5 vonalakhoz. Létrehoztunk két recens népességet jellemző adatbázist, 102 anyaországi „magyar” és 76 „székely” egyén mitokondriális DNS-adataival. Az egyes haplocsoport-megoszlások azt mutatták, hogy a két recens mintában az európai típusú haplocsoportok dominanciája figyelhető meg: 99 % a mai magyar népesség és 97,4 % a székely populáció körében. A típusok eloszlása nagyon hasonló képet mutatott a többi közép-európai populáció haplocsoport-eloszlásához, de az ázsiai típusú genetikai elemek kis százalékban mind a magyar, mind a székely populációban jelen vannak (M, illetve C haplocsoportok) (5a. ábra). A rendelkezésünkre álló szekvenciákat statisztikai analízisnek is alávetettük. A 10. századi szekvenciákat összehasonlítottuk 73 európai, illetve ázsiai populációból származó szekvenciákkal, beleértve mai „magyar” illetve „székely” populációkból származó mintákat is. A közel 10 500 szekvenciát magába foglaló analízis során genetikai távolságot számoltunk az egyes populációk között, majd ezeknek kétdimenziós ábrázolásával jelenítettük meg a populációk közötti rokonsági kapcsolatokat (6. ábra). A kétdimenziós ábrázolás abszcisszája mentén egyértelműen kivehető a populációk kelet-nyugati irányú elhelyezkedése. A 10. századi minták egy csoportban térképeződnek a „török”, „kurd”, „ukrán”, „palesztin”, „szír”, „iraki”, „észak-oszét”, „azeri” „komi” populációkkal. Ez a csoport egy közép-ázsiai populációkat magába foglaló kisebb, illetve a nyugat-európai populációkat magába foglaló nagyobb csoport között helyezkedik el. A későbbiekben tárgyalt hartai temető szekvenciáit külön értékelve a genetikai távolságok alapján is egy keleti (ázsiai) irányú eltolódás figyelhető meg. A hartai populáció az összes honfoglaló mintát is magába foglaló csoport és a belső-ázsiai populációk között helyezkedik el, vagyis abban a genetikai távolságok alapján még kifejezettebb ázsiai hatás érvényesül. Az anyai ági rokonság eddigi vizsgálata alapján tehát elmondható, hogy a honfoglaló populációban európai és ázsiai típusú mitokondriális genetikai elemek is előfordulnak, de jelentősen több ázsiai típusú elemet tartalmazva, mint a recens „magyar” és „székely” minták. Utóbbiak ugyanakkor a jelenkori európai népességekre jellemző genetikai képet mutatnak. Jelenlegi adataink arra nem nyújtanak lehetőséget, hogy e különbség népességtörténeti hátterét kronológiai viszonylatban is értékeljük, hiszen egyelőre nem rendelkezünk mtDNS-adatokkal a Kárpát-medencének sem a 10. századot követő, sem azt megelőző évszázadairól.
2002-ben a pályázat keretében feltárt, huszonegy egyén, főként nők maradványait rejtő klasszikus honfoglalás kori leletanyagú hartai temető vizsgálata több szempontból is meglepő eredményekkel szolgált. A csontok jó DNS-megőrződésének köszönhetően tizenkilenc vázból vett mintából sikerült a haplokategorizálás, valamint a besorolás megerősítése további mutációs pontok meghatározásával, hét esetben sikerült a genetikai nem meghatározása is. Az eltérő anyai vonalak jelenléte a temetőben a vártnál jóval magasabb volt, legalább tizenegy család jelenlétét igazoltuk (H, U, U5a, U8b, JT, T3, V, N1a, A, C és D). A heterogén megoszlás miatt csupán három minta mutációs mintázata mutatott rokonságot, vagyis ezeknél nem zárható ki az egyenes anyai ági leszármazás. A régészeti prekoncepcióból kiindulva azt vártuk volna, hogy a genetikai nyalábok a temetőt használó népességről egy sokkal egyöntetűbb képet rajzolnak ki, és a nők magas száma a temetőben (84 %) szintén egy homogénebb anyai ági vonalat valószínűsített. Az a tény, hogy a vizsgált egyének nem állnak genetikailag rokonságban egymással, arra enged következtetni, hogy a közösség vagy nem vérségi (családi) rendben szerveződött, vagy a temető használati ideje nem tette lehetővé, hogy a családi kapcsolatrendszer a temetkezésekben is leképeződjön. Erre utalhat, hogy a temetőben egyetlen kisgyermeknek sem találtuk meg az anyai felmenőjét! Meglepő eredmény, hogy az eddig megvizsgált 10. századi temetők népességéhez képest a hartai temető csontanyagában kimagaslóan magas az ázsiai haplocsoportok előfordulása. E temető vizsgálata elsősorban a régészet számára hozhat áttörést, kikényszerítve új módszertani szempontrendszer(ek) kialakítását. A temetőt kezdetben a lelőhelyen talált viszonylag kevés sír miatt ún. „kiscsaládi temetkezési rendszer” keretei közt értelmeztük, de a genetikai eredmények ellentmondtak ennek. A régészeti kutatás általában – alapvetően néprajzi szempontok, de a történeti források alapján is – a kora középkori temetőket családi rendszerben képzelte el, a kutatók a „családi genealógiát” még olyan esetben is rávetítették a lelőhelyekre, ahol arról nyilvánvalóan nem lehetett szó. Vizsgálati eredményünk alapjaiban rendítette meg ezt a modellt. A kiscsaládi temetőmodell befolyásolta a kronológiai szemléletet is, hiszen ezáltal a korábban a családon belüli születési sorrendből levezetett tipokronológia új, lényegesen rövidebb periódusokra osztható megközelítéssel számolt. A vizsgálati eredményt jelenleg kétféle módon tudjuk interpretálni: 1. a női sírok „feleségek”, „ágyasok” maradványait rejtik; 2. a temető nem családi rend alapján alakult ki. Mindkét értelmezés magában hordozza a következtetést, hogy a temető használati ideje igen rövid lehetett. A hartai temető példája azt is mutatja, hogy egy lokális mintavételi sorozat (teljes temető) mitokondriális mintázata jelentősen eltérhet különböző temetkezési helyek összesített adatbázisától.
Populációk apai ági leszármazási vonalát az Y-kromoszóma azon szakaszainak vizsgálatával követhetjük nyomon, melyek polimorfak, és a meiózis során nem rekombinálódnak. Az Y-kromoszóma generációról generációra viszonylag változatlan formában öröklődik az apai vonalon. Jelentős részén csak mutációval jöhet létre változás, nem keveredik az anyai és az apai információ. Az Y-kromoszóma legfontosabb biológiai szerepe a nem meghatározása, és a férfi fertilitás biztosítása. Haploid, vagyis sejtenként csak egy kópiában fordul elő. Az Y-kromoszóma mutációiból végigkövethetjük a férfiak genetikai történetét, amely földrajzi és filogenetikai értelemben nagyjából megegyezik azzal, amit a mitokondriális DNS-ből kiolvashatunk. Az Y-kromoszomális haplocsoportokat a biallélikus markerek határozzák meg, melyek lehetnek pontmutációk (ún. SNP – Single Nucleotide Polymorphism – markerek), ill. inszerciós és deléciós polimorfizmusok. Vizsgálatukkal átlátható módon végig tudjuk vezetni az anatómiailag modern ember vándorlásainak idejét és irányát, illetve következtethetünk a különböző populációk apai vonalának geográfiai eredetére, más népcsoportokkal való keveredésére, rokonsági viszonyaira. Különös jelentőséggel bír számunkra a Tat polimorfizmus (T>C) derivált C allélja, mely az N3 haplocsoportot határozza meg. Ez a haplocsoport széles körben elterjedt Európa északi és keleti területein, ugyanakkor gyakorlatilag nem fordul elő a kontinens nyugati és déli részein élő populációkban. Ázsiában az északi területeken élő népcsoportokban fordul elő számottevő gyakorisággal. A szakirodalom alapján a Tat C allél jelen van a legtöbb uráli nyelvet beszélő populációban, Y-kromoszomális génkészletüknek jelentős részét képezve, ugyanakkor a magyarul beszélő népcsoportok esetében ez a mutáció szinte egyáltalán nem fordul elő. Az általunk vizsgált modern „magyar” (132 fő) és „székely” (97 fő) férfiak közül csupán egyetlen „székely” férfiban volt jelen a Tat C allél. E megfigyelések alapján felvetődik a kérdés, hogy a honfoglalás kori populációban jelen volt-e ez a mutáció vagy sem. Az eddigi munkánk során hét csontlelet esetében végeztük el a tipizálást. A vizsgált minták közül két esetben a mutáns C allél jelenlétét, míg öt leletnél a vad típusú T allél előfordulását igazoltuk (1. táblázat). A sikeresen vizsgált esetszám igen alacsony, azonban a vizsgálatokat az archaikus DNS-minták esetében megnehezíti, hogy az ásatag csontokban épen maradt DNS-szakaszok mennyisége limitált, ugyanakkor az Y-kromoszóma minden hímnemű egyén sejtjében csak egy példányban van jelen. Eredményeink alapján a Tat C allél jelen volt a honfoglalás kori populációban, illetve a Tat polimorfizmus megfelelő genetikai bélyeg a 10. századi populáció Y-kromoszomális genetikai összetételének analíziséhez. A C allél jelenléte egyben alátámasztja eredményeink hitelességét is, hiszen sem kutatócsoportunk férfi tagjai, sem a Régészeti Intézet férfi munkatársai (régész, antropológus) – akik kapcsolatba kerülhettek a csontmintákkal – nem hordozzák ezt a mutációt.
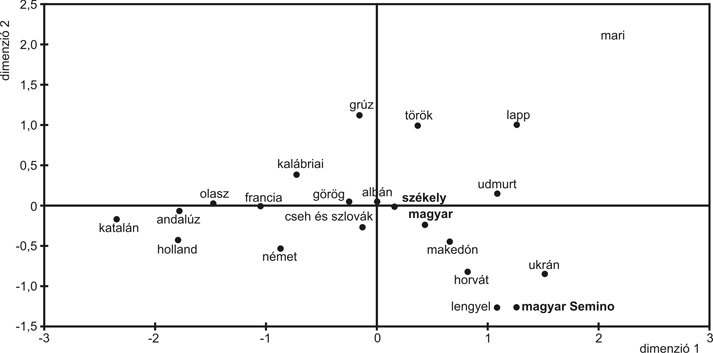
A recens „magyar” és „székely” populációban, összesen 197 egyénnél, huszonkét Y-kromoszomális biallélikus markert tipizáltunk. A vizsgált „magyar” férfiak (100 fő) Magyarország különböző területein születtek, de a leginkább reprezentált régió az Alföld területe volt (100/90). A „székely” minták (97 fő) Korondról, Erdély területéről származnak. Minden minta esetében először az M96, az M89, az M9 és az M45 SNP markereket analizáltuk, így minden egyes mintát el tudtunk helyezni a filogenetikai fa valamelyik nagyobb ágán. A markerek hierarchikus szerveződésének megfelelően tudtuk, hogy milyen további markereket kell még vizsgálnunk a pontos kategorizálás érdekében. Ez utóbbi tizennyolc marker (M35, M78, M170, M253, P37, M26, M201, P15, M304, M267, M172, M102, M67, M92, Tat, M173, M17, M269) allélikus állapotát a PCR-reakciót követően restrikciós emésztéssel határoztuk meg. A vizsgált markerek segítségével a modern magyar és székely mintákat összesen tizenhét Y-kromoszomális haplocsoportba soroltuk be (7. ábra). A megfigyelt haplocsoportok általánosan jellemzik a különböző európai populációkat, egyetlen kivételt képez ezalól a Közép-Ázsiában gyakori P*-M45 (nem-M173) klaszter , amely a vizsgált „székely” populáció 3,1 %-át jellemzi. Mindkét populációban az M173-as marker által meghatározott R1 haplocsoport fordul elő a legnagyobb gyakorisággal („magyar”: 45 %, „székely”: 38,2 %). Ez a marker kb. 30–35 ezer éve terjedt el Európában Közép-Ázsia irányából, és a mai európai férfiak mintegy 50 %-át jellemzi. A markert hordozó európai férfiak alapvetően két, Európában egymással ellentétes földrajzi megoszlást mutató szubkládba, a R1a1 és az R1B3 haplocsoportba sorolhatók be. Az R1a1 csoport Kelet-Európában gyakori (30–60 %), illetve Közép-Ázsiában és India északnyugati részén. A vizsgált „magyar” populációban 30 %-os értékkel ez a leggyakoribb klaszter, míg a „székely” mintában 18,6 %-os gyakorisággal van jelen. A haplocsoport elterjedésében legalább három folyamat játszhatott szerepet: 1. Európa rekolonizációja a jégtakaró visszahúzódása után a mai Ukrajna területén lévő refúgiumból, ahol valószínűleg ez a mutáció kialakult; 2. vándorlások az északi Pontus-vidék területéről Kr. e. 3000– 1000; 3. a szláv migráció Kr. u. 5–7. sz. Az R1b3 haplocsoport, bár egész Nyugat-Eurázsiában elterjedt, a legnagyobb gyakorisággal (40–80 %) Nyugat-Európában fordul elő. A „magyar” férfiak 15 %-a, a „székely” férfiak 19,6 %-a tartozik ebbe a klaszterbe. Az irodalmi adatok szerint a haplocsoport (az ezt definiáló M269-es marker kb. 10–15 ezer éve alakult ki, valószínűleg az Ibériai-félsziget/Délnyugat-Franciaország területén) elterjedési mintázata az európai kontinens Ibéria, ill. Kis-Ázsia területéről való benépesítését tükrözi a felső paleolitikum és a holocén idején. Az I haplocsoport hasonló gyakorisággal fordul elő a magyarul beszélő populációkban („magyar”: 24 %, „székely”: 21,7 %). Ez a klaszter az egyetlen olyan Y-kromoszomális haplocsoport, amely szinte kizárólag csak az európai kontinensen van jelen. A haplocsoportot definiáló M170-es marker kb. 22 ezer éve alakulhatott ki Európában, és a mai európai Y-kromoszomális génkészlet kb. 20 %-át jellemzi. A két fő szubkládja közül az I1a Észak-Európában gyakori, míg az I1b Kelet-Európában és a Balkánon elterjedt. Közép-és Kelet-Európában az I1a és az I1b haplocsoport átfedő gyakorisági grádienst mutat. Ez a két leszármazási vonal ellentétes megoszlási mintázatot mutat az általunk vizsgált „magyar” és „székely” populációban. Míg ugyanis az I1b haplocsoport előfordulási gyakorisága több mint kétszeres a „magyar” férfiak esetében a „székely” mintához viszonyítva, addig az I1a kétszeres mértékben fordul elő a „székely” populációban a „magyar” csoporthoz képest. Elképzelhető, hogy az I1a, illetve a fentebb említett R1b3 haplocsoport magasabb gyakorisága a „székely” populációban a 12. századtól Erdély területére betelepülő germán eredetű szászok genetikai hatását tükrözi. A J, az E3b és a G haplocsoportok európai elterjedését a neolitikum idején a Közel-Keletről érkező földművelő népek expanziójához kötötték. A J és E haplocsoportok újabb filogeográfiai analízisének eredményei azt mutatják, hogy a neolitikumot követően még számos migrációs esemény járult hozzá ezeknek a leszármazási vonalaknak az elterjedéséhez. A J klasztert definiáló polimorfizmus a Közel-Keleten alakult ki, gyakorisága a Közel-Kelet irányából Európa felé csökken. Európában a déli-délkeleti területeken (a mediterrán területek középső régiójáig) fordul elő nagyobb gyakorisággal (>20 %), viszont ritkán fordul elő a Balkán északi részén, illetve Közép-Európában. A J haplocsoporton belül a J1 és a J2 klasztert különböztethetjük meg. A J1 haplocsoport nagy gyakorisággal (10–30 %) van jelen a Közel-Keleten, Észak-Afrikában és Etiópiában, míg az európai területeken ritkán fordul elő (0–6 %). Elterjedésében fontos szerepet tulajdonítanak az arab diffuziónak. Míg a „magyar” férfiaknál talált 3 %-os gyakorisági érték hasonló a Közép- és Kelet-Európában tapasztalthoz, addig a „székely” mintában kapott érték (10,3%) ennél magasabb. A J2 klaszter sokkal elterjedtebb Európában, szubkládjai közül a J2e és a J2f* Európában és Ázsiában is előfordul. A J2e haplocsoportot szinte teljes mértékben a J2e1 szubklaszter reprezentálja, amely valószínűleg a Balkán déli részéről terjedt el Európa többi területére. A J2f* haplocsoport a Kaukázus területén élő populációkban fordul elő a leggyakrabban, míg a J2f1 elsősorban Törökországtól nyugatra van jelen, a mediterrán területek északi részén. Európában a J haplocsoporton belüli diverzitás valószínűleg Törökország és az Égeikum területére vezethető vissza. Vizsgálatainkban a J haplocsoport meglepően gyakori előfordulását tapasztaltuk a „magyar és a „székely” (16 %, illetve 21,6 %) populációban. A korábbi adatok szerint a J haplocsoport ritkán, csak 2–3 %-ban fordul elő a magyar nyelvű populációkban, míg egy másik kutatócsoport által tipizált „magyar” csoportban 14 %-os gyakorisággal volt jelen. Valószínűleg a mintavételi eltérés eredményezte, hogy az általunk nyert gyakorisági adatok eltérnek a korábbi irodalmi értékektől. Míg az általunk tipizált „magyar” minták elsősorban az Alföld területéről származnak, addig például Ornella Semino és munkatársai Budapestről, Egerből és Mátraderecskéről gyűjtöttek mintákat, utóbbi területeken elsősorban palócok közül. A J haplocsoport nagyobb gyakorisága az általunk vizsgált populációkban egyrészt utalhat anatóliai, illetve dél-balkáni genetikai hatásra, ugyanakkor genetikai sodródás következménye is lehet, főként a zártabb közösségekben élő székelyek esetében. A J klaszter magasabb értéke összefügghet az Ottomán Birodalom terjeszkedésével, illetve ennek hatására a balkáni régióban élő népességek Kárpát-medencébe településével. Elképzelhető, hogy az ősi magyar populációnak is szerepe lehetett a J haplocsoportnak a Kárpát-medencében való elterjesztésében. A mitokondriális DNS vizsgálataink azt mutatták, hogy mindazok a populációk, melyek kapcsolatba kerültek a korai magyar népcsoporttal, nyomot hagytak azok génkészletében. A mtDNS-alapon végzett statisztikai analízis azt mutatta, hogy az ősi magyar populáció közeli genetikai távolságra helyezkedik el nemcsak néhány közép-ázsiai és európai népcsoporttól, hanem néhány közel-keleti populációtól is. Utóbbiakban pedig a J haplocsoport nagy gyakorisággal van jelen. Az E3b haplocsoport hasonló gyakorisággal fordul elő a vizsgált „magyar” és a „székely” populációban (10 %, illetve 9,2 %). Az E3b1 haplocsoport széles körben elterjedt, jelen van Észak- és Kelet-Afrikában, a Közel-Keleten és Európában is, ahol a leggyakrabban előforduló E3b szubklasztert reprezentálja. A haplocsoport elterjedésének iránya Európa déli, délkeleti része felöl a Vardar–Morava–Duna vonala lehetett kb. 7300 évvel ezelőtt. Az E3b leszármazási vonalak jelenlegi európai megoszlási mintázatának kialakításában fontos szerepe lehetett a kb. 7–8000 éve a Balkán területéről a Nyugat-Európa és Közép-Európa déli része felé irányuló expanziónak. Eredményeinket statisztikai alapon is kiértékeltük. A kapott távolság-mátrix értékeit egy kétdimenziós térben ábrázoltuk, ahol az egyes populációknak megfelelő pontok közötti lineáris távolság egyenesen arányos a köztük lévő genetikai távolsággal (8. ábra).
A statisztikai vizsgálat szerint az általunk vizsgált „magyar” és
„székely” populáció genetikailag közeli rokonságban áll egymással, illetve más
közép-európai (például: cseh és szlovák) és balkáni populációkkal. Ugyanakkor
egyértelműen elkülönül egymástól az
általunk analizált és a Semino és munkatársai által tipizált magyar csoport. Ez
utóbbi közel térképeződik a lengyel és
az ukrán populációhoz. Az eltérésre lehetséges magyarázat, hogy a Semino-féle
kutatócsoport elsősorban
észak-magyarországi mintákat vizsgált, ahol jelentős volt a szomszédos északi területek más
anyanyelvű népességeivel való
diffúzió. Valószínűleg ez az esemény
felelős az általuk tapasztalt kiugróan
magas R1a1 haplocsoport gyakoriságért is (60 %, míg az általunk vizsgált
mintánál 30 %).
IRODALOM Csányi Bernadett et al. (2008): Y-chromosome Analysis of Ancient Hungarian and Two Modern Hungarian-Speaking Populations from the Carpathian Basin. Annals of Human Genetics. 72, 519–534. Cann, R. L. – Stoneking, M. – Wilson, A. C. (1987): Mitochondrial DNA and Human Evolution. Nature, 325, 31–36. Cruciani, Fulvio et al. (2002): A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes. American Journal of Human Genetics. 70, 1197–1214. Di Giacomo, F. et al. (2004): Y Chromosomal Haplogroup J As a Signature of the Post-Neolithic Colonization of Europe. Human Genetics. 115, 357–371. Giles, R. E. – Blanc, H. – Cann, H. M. – Wallace, D. C. (1980): Maternal Inheritance of Human Mitochondrial DNA. Proceedings of the National Academy of Sciences of the USA. 77, 6715–6719. Jobling, M. A. – Tyler-Smith, C. (2003): The Human Y Chromosome: An Evolutionary Marker Comes of Age. Nature Reviews Genetics. 4 Aug. 8, 598–612. Kristó Gyula (2002): A székelyek eredete. Balassi, Bp. Maca-Meyer, Nicole et al. (2001): Major Genomic Mitochondrial Lineages Delineate Early Human Expansions. BMC Genetics. 2, 13. Pálffy Géza (2000): A tizenhatodik század története. Pannonica, Budapest http://www.3d.hu/hkkk/files /tanacsadok/palffy_16sz_tortenete.pdf Peričć, Marijana et al. (2005): High-resolution Phylogenetic Analysis of Southeastern Europe Traces Major Episodes of Paternal Gene Flow among Slavic Populations. Molecular Biology and Evolution. 22, 10, 1964–1975. Richards, Martin et al. (1998): Phylogeography of Mitochondrial DNA in Western Europe. Annals of Human Genetics. 62, 241–260. Richards, Martin et al. (2000): Tracing European Founder Lineages in the Near Eastern mtDNA Pool. American Journal of Human Genetics. 67, 1251–1276. Semino, Ornella et al. (2000): The Genetic Legacy of Paleolithic Homo Sapiens Sapiens in Extant Europeans: A Y Chromosome Perspective. Science. 290, 1155–1159. Semino, Ornella et al. (2000): MtDNA and Y Chromosome Polymorphisms in Hungary: Inferences from the Palaeolithic, Neolithic and Uralic Influences on the Modern Hungarian Gene Pool. The European Journal of Human Genetics. 8, 5, 339–346. Tömöry Gyöngyvér et al. (2007): Comparison of Maternal Lineage and Biogeographic Analyses of Ancient and Modern Hungarian Populations. American Journal of Physicical Anthropology. 134, 3, 354–368. Underhill, Peter A. et al. (2000): Y Chromosome Sequence Variation and the History of Human Populations. Nature Genetics. 26, 358–361. Wells, Spencer et al. (2001): The Eurasian Heartland: A Continental Perspective on Y-Chromosome Diversity. Proceedings of the National Academy of Sciences of the USA. 98, 18, 10244–10249. Zerjal, T. et al. (1997): Genetic Relationships of Asians and Northern Europeans, Revealed by Y-Chromosomal DNA Analysis. American Journal of Human Genetics. 60, 1174–1183.
1. ábra • Az emberi örökítőanyag megoszlása a sejten belül
2. ábra • Az archaikus minták vizsgálatának menete
3. ábra • Az európai haplocsoportok kialakulása
1. táblázat • A mintavétel és a haplokategorizálás adatai – *egy
mintát mindkét szakaszban vizsgáltunk, ezért a végső adat 68 db. sikeresen kategorizált
minta
4. ábra • A vizsgált 10. századi minták haplocsoport-megoszlása
5. ábra • Recens magyar nyelvű populációk haplocsoport-megoszlása
6. ábra • A populációk közötti genetikai távolság kétdimenziós ábrázolása – 1: iraki, 2: szír, 3: palesztín, 4: örmény, 5: azeri, 6: észak-oszét, 7: bolgár, 8: román, 9: albán, 10: olasz, 11: szicíliai, 12: szardiniai, 13: észak-portugál, 14: közép-portugál, 15: dél-portugál, 16: észak-spanyol, 17: közép- és dél-spanyol, 18: galíciai, 19: baszk, 20: svájci, 21: osztrák, 22: lengyel, 23: orosz, 24: cseh, 25: dán, 26: svéd, 27: norvég, 28: izlandi, 29: cornwalli, 30: walesi, 31: skót, 32: észt, 33: karéli, 34: belga, 35: angol, 36: német, 37: ír, 38: kurd, 39: francia, 40: breton, 41: adige (cserkesz), 42: fehérorosz, 43: Bosco Gurin-i, 44: bosnyák, 45: burját, 46: kanári, 47: horvát, 48: európai-kaukázusi, 49: venki, 50: finn, 51: grúz, 52: görög, 53: krétai, 54: kazak, 55: kirgiz-magasföldi, 56: kirgiz-alföldi, 57: komi, 58: mari, 59: moksa, 60: mongol, 61: oberwallisi, 62: oszét, 63: rétoromán, 64: lapp, 65: szerb, 66: szlavón, 67: szlovák, 68: török, 69: ujgur, 70: ukrán, 71: csengelei kun minták 72: mai magyar, 73: székely, 74: 10. századi honfoglaló minták, 75: hartai honfoglalás kori minták
2. táblázat
• A vizsgált minták allélikus
állapota és lelőhelye
7. ábra • A vizsgált modern „magyar” és „székely” populációban előforduló haplocsoportok genealógiája és előfordulási gyakorisága
8. ábra • Kétdimenziós MDS-grafikon genetikai távolság alapján
A cikk eredetioldala: http://www.matud.iif.hu/08okt/05.html
|